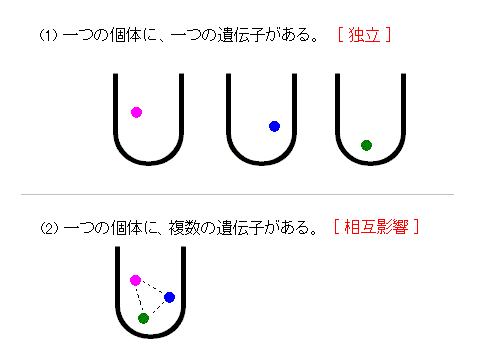
(1) では、それぞれの遺伝子は、独立している。たがいに相互的な影響がない。それぞれの遺伝子の有利・不利だけを、独立的に考えればいい。
(2) では、それぞれの遺伝子は、関連している。たがいに相互的な影響がある。それぞれの遺伝子の有利・不利だけでなく、他の遺伝子からの影響をも、考慮する必要がある。
以下の文章のうち、ドーキンス(利己的遺伝子説)について言及した分は、内容が古くなっています。
核心を突いた新たな内容は、次の文書をご覧ください。
→ 利己的遺伝子とは
→ 血縁淘汰説とは
重要な点は、前者の文書の、前半部分に書いてあります。
詳細は、後者の文書に、要点が記してあります。細かな点は、そこからリンク先をたどってください。
なお、本文書(現在見ているこの文書)は、利己的遺伝子説についてはピンボケな点(核心でない点)が多くありますが、それぞれの論点は、進化論の話題の一部として、読むことができます。(ただし、あまり重要な論点ではありません。本文書の全体は、特に重要な話題を述べていません。)
【 要点 】
この文書の要点を示す。
私の主張について、「ドーキンス説を全否定している」という見解がある。だが、それは誤解である。まず、大方の見解は、こうだ。ドーキンス説は、「増えるべき遺伝子が増える」というようなことを言っているだけである。当り前のことを言っているだけだ。ゆえに、ドーキンス説は特に否定するには値しない。(つまり、ドーキンスは言い換えをしただけであって、学問的な業績は何もない。)ここから、「ドーキンス説を否定する南堂久史は(トートロジーを否定しているから)トンデモだ」という見解が生じる。だが、これは大いなる誤解である。
私がドーキンス説を否定しているのは、上記の点ではない。以前の説に対するドーキンス説の独自性を否定しているのではない。むしろ、以前の説もひっくるめて、これまでの進化の原理を否定している。すなわち、次の説だ。
「有利な遺伝子(増えるべき遺伝子)は増える。だから、進化が起こる」
私は、緑の部分を否定しているのではない。紫の部分を否定しているのだ。(多くの人はそこを誤解する。)
ここで、緑の部分は、小進化の説明だ。紫の部分は、大進化の説明だ。
この文の全体を見ると、緑の部分(≒ 小進化)は成立するが、紫の部分(≒ 大進化)は成立しない。つまり、小進化の蓄積が大進化になるわけではない。──それがつまり、私の説だ。
私は、ドーキンス説に固有な特徴を否定しているのではなく、大進化の原理そのものを否定している。私が何を否定しているか、勘違いしないでほしい。
「南堂久史はドーキンス説を読んでいない」という批判をよく聞くが、彼らの信じているドーキンス説(緑の部分)と、私の指摘しているドーキンス説(紫の部分)とは、言葉はどちらも「ドーキンス説」であっても、意味するところが異なる。
また、適用場面も、私は大進化だけに言及しているのであって、小進化には言及していない。この点も誤解しないようにしてほしい。
( ※ なお、南堂説とは何かというと、次のことだ。
「大進化は、小進化の蓄積ではない。」
「大進化には、小進化とは別の原理がある。」
この主張のもとで、新たな進化の原理を提出する。それが南堂説。)
|
遺伝子のうち、育児をしたがる遺伝子を持つ個体は、育児をするので、その結果として、その個体の子孫の細胞に含まれる同種の遺伝子を繁殖させることができるので、結果的にその遺伝子が多数派になる。ここでは、育児をするという遺伝子があって、そういう遺伝子が親である個体を操作した、というふうになる。つまり、「遺伝子が個体を操作する」というふうになる。
(Openブログの読者コメントからの引用。)
遺伝子のうち、その遺伝子を持つ個体の脳をして「自分の子がかわいい」と思わしめるような脳を構成することに成功した遺伝子は、自分を細胞内に持つ個体が、まさに自分の子をかわいいと思うがゆえに、自分の子を安全に育成し、子孫を残しやすくなるので、その結果として、その個体の子孫の細胞に含まれる「自分と大変よく似た遺伝子」を繁殖させることができるので、結果的に遺伝子としての多数派になる。ここでは、「わが子はかわいい」と思うような遺伝子があって、そういう遺伝子が個体を操作した、というふうになる。つまり、「遺伝子が個体を操作する」というふうになる。
(Openブログの読者コメントからの引用。)
|
|
(1) では、それぞれの遺伝子は、独立している。たがいに相互的な影響がない。それぞれの遺伝子の有利・不利だけを、独立的に考えればいい。 (2) では、それぞれの遺伝子は、関連している。たがいに相互的な影響がある。それぞれの遺伝子の有利・不利だけでなく、他の遺伝子からの影響をも、考慮する必要がある。 |
【 注記 】
ここで「浮気」というのは、通俗的な例である。話をわかりやすくするために、簡易的に比喩的に表現した。現実には「浮気の遺伝子」という単独の遺伝子はない。
学術的に言うなら、「ミツバチが自分の妹を育てるという本能の遺伝子」という例を取る方がいい。ただし、そういう言い方だと、厳密ではあるが、頭に入りにくい。「浮気の遺伝子」という話の方が、頭にすぐ入る。
だからここではあえて、不正確ではあるが「浮気の遺伝子」という言葉で説明した。学問的に考えるときには、「ミツバチが自分の妹を育てるという本能の遺伝子」というふうに、いちいち置き換えて読み直してほしい。
( ※ 「そんなことは、いちいち言われなくてもわかっている」と思う人が多いだろう。しかし世の中、読解力のない人が多いので、ここに念のために注記しておいた。)
|
表紙ページ http://hp.vector.co.jp/authors/VA011700/biology/